Human and non-human communication
A curious controversy
Homo sapiens may be the only genuinely musical species.
At least, no other species seems to have songs in which musical intervals (small whole-number ratios between pitches) play a role. No other species seems to create rhythmic patterns by subdivisions of a beat.
There might even be some related evolutionary specialization of the auditory system -- at least, the just noticeable difference in pitch perception is about an order of magnitude smaller for humans than for other mammals.
Then again, maybe this is going too far. What about the songs of whales or songbirds? Don't they have real melodies? Don't apes sometimes beat out rhythms?
The point is, no one seems to get worked up about this question. Books and articles are not written about whether the songs of whales or sparrows are Truly Music. There is plenty of study, scientific and otherwise, of the vocal displays of whales and songbirds. However, the interest is in what animal songs are like -- what they sound like, how they are made, how they are learned, what they are used for -- and not whether they basically the same as, or basically different from, human music.
By comparison, there is enormous and continuing controversy about whether various sorts of animal behavior, both natural and taught by human trainers, are Truly Language.
Why?
For some reason, people find it more interesting -- either more attractive or more threatening -- to think of animals as having command of language, than to think of them as having command of music. The debate often seems to take on a quasi-religious tone, as if the issue were whether animals had souls rather than whether they have languages.
As with many such controversies, it's hard to stay out of the fight. Nevertheless, we are going to try, in part by staying away from the vexed case of apes and sign language, and focusing on natural communication among animals.
What natural animal communication is like
 When a dog -- or
a wolf or a coyote -- lowers its forelegs to the ground and waves its tail,
this is a way of saying, "everything that follows is just a game. Are you
ready to play?" Ethologists (students of animal
behavior) call this the Canid Play Bow. Cats apparently don't
understand this bit of dog metalanguage, though most dog owners do.
When a dog -- or
a wolf or a coyote -- lowers its forelegs to the ground and waves its tail,
this is a way of saying, "everything that follows is just a game. Are you
ready to play?" Ethologists (students of animal
behavior) call this the Canid Play Bow. Cats apparently don't
understand this bit of dog metalanguage, though most dog owners do.
The Canid Play Bow is an example of what ethologists call a display, meaning roughly a salient pattern of behavior.
Displays are modulated in intensity or clarity of presentation, with similarly gradient implications for their "meaning." For instance, the eastern kingbird has a vocalization said to sound like "zeer" that indicates possible aggressive intent. The sharpness and harshness of the call varies, and the probability that the animal will actually attack increases as the sharpness/harshness of the call increases.
Sometimes displays are quite specific and complex in their behavioral structure and also in their apparent function. Other displays have a specific form, but their function -- or meaning -- that is more difficult to define. For instance, the blacktailed prairie dog has a display called the "Jump-Yip," which involves throwing the forepaws into the air, pointing the nose straight up, and emitting an abrupt two-part vocalization. The Jump-Yip seems to mean "I'm trying to decide whether to run away or do something else that might be more important to me." A bit less than half the time, a Jump-Yip is followed by flight, but there always seems to be an alternative: to make sexual advances to another prairie dog, to challenge a neighbor aggressively, to take a dust bath, or whatever. The display is only used when there is an alternative to flight, but the alternative could be just about anything.
More often, displays are fragments of behavior that may be combined in
various ways, and may shade gradually into what may be considered an entirely
different display. For example, here is one ethologist's version of the
system of dogs' body language, of which the "play bow" is one part:
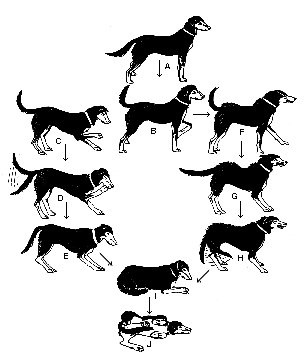 Changes
in body postures and facial expressions by which dogs indicate their feelings
and intentions to other dogs.
Changes
in body postures and facial expressions by which dogs indicate their feelings
and intentions to other dogs.
-
A-B: Neutral to alert attentive positions
C: Play-soliciting bow
D-E: Active and passive submissive greeting -- tail wags, ears fold back, weight is transferred to hind legs
I: Passive submission
J: Rolling over and showing belly and genitals
F-H: Gradual shift from aggressive display to ambivalent fear/defensive/aggressive posture
There are other cases, for instance warning calls, where displays may have a range of fairly specific referential content. For instance, vervet monkeys give one kind of alarm call when they see a snake, another kind when they see a leopard, and a third kind when they see an eagle. This set of referential distinctions is useful, because different kinds of defensive behavior are appropriate in the three cases.
In response to the snake call, the troupe of vervets will all stand up on their hind legs in the open and look around on the ground -- to find the snake. If the predator were an eagle or a leopard, standing up in the open would be very unwise.
In response to the leopard warning call, the members of the troupe run up to the top of the nearest tree, where the heavy leopard can't follow them. This would be just the wrong thing to do to avoid an eagle, and not optimal for a snake either.
In response to the eagle warning call, the members of the troupe run
into a nearby bush or under the lower branches of a nearby tree. Since
this is where leopards hide while stalking, and is as likely as anywhere
to be on top of a snake, this behavior is again a good response only to
the type of predator that the particular call signals.
Types of display
Just about anything that can be controlled can become part of a "display". Electric eels and fish use electromagnetic signaling in courtship and establishment of territory. Cuttlefish and other cephalopods have chromatophores and other specialized cells in their skin that allow them to change its color and texture in complex patterns. In one species, observers have catalogued 31 distinct full-body patterns with behavioral significance. For instance, male cuttlefish flash a zebra-striped pattern in circumstances appropriate for mating. Females generally remain mottled, but if a female turns a uniform gray, it signals that she is ready to mate. The dominant male will approach the female and also turn a uniform gray. If another male approaches, the male involved in the mating will turn on zebra stripes on the side facing the potential rival, while retaining his sexy gray color on the side facing the female.
Communicative functions in animals
We've already seen some of the functions of animal displays, vocal or otherwise: warning of predators, attracting mates or scaring off sexual rivals, signaling aggressive or submissive attitudes.
We can try to make a more complete list of functions. Here is one, adapting from a text on the topic:
- To advertise individual identity, presence, and behavioral predispositions.
- To establish social hierarchies without the disruption of actual fighting or fleeing.
- To synchronize the physiological states of a group, e.g. for breeding.
- To monitor the environment collectively for dangers and opportunities
- To synchronize organized activities (hunting or foraging, migration, etc.)
- To discourage predators (e.g. extravagant leaping and kicking by some antelopes)
- To lure prey (e.g. predatory mimicry by fireflies)
Evolution of animal communication
Most animal displays are thought to develop by a process of ritualization of previously existing behavior. A selective advantage may come to exist for those individuals who use certain behaviors in a way that is partly different from the norm, thereby changing the norm in a certain direction. For instance, urine tends to smell; this can lead to specialized use to mark territory, or develop into chemical sex attractants, resulting in the development of urination behaviors that have nothing to do with the need to void wastes.
Other displays develop from ritualization of intention movements,
i.e. incomplete acts, such as turning away or starting to fly. A related
example is the development of frowning from protective eyebrow-lowering,
which presumably was originally part of more general preparations for
attack or defense. Movements may be redirected or displaced from their
normal context, repeated unnecessarily, etc. as part of ritualization.
Instinctive vs. learned patterns
The patterns of animal communication seem to be largely, though not entirely, genetically determined.All vervet monkeys use the same set of warning cries. However, the particular creatures that evoke them vary from place to place, depending on the local types of predators. Baby vervets start out pretty undiscriminating, giving "eagle" warning calls to any reasonably large bird, including those that are completely harmless, and "leopard" warning calls to any largish terrestrial animal, including grazing animals, anteaters etc. They gradually learn -- apparently from the reactions of their elders -- how not to 'cry wolf.'
Most animals learn to respond appropriately to the warning cries of other species that happen to live near them.
Some species -- for instance some songbirds -- have "dialects", in which some properties of their vocalizations or other displays are learned, during a critical period early in each animal's life. The resulting pattern of geographical variation in the vocal displays then plays a role in social structure -- just like human shibboleths. It is said that such dialect variation tends to occur (in songbirds) just in case the species colonizes a wide range of microhabitats, where each local group becomes especially adapted to their local conditions, and will benefit from breeding within the group to maintain the adaptations intact.
 A more elaborate
form of social evolution occurs in the songs of humpback whales. Male
humpbacks "sing" long (half-hour) and complex "songs", apparently as part
of lek behavior. also called arena behavior.
A more elaborate
form of social evolution occurs in the songs of humpback whales. Male
humpbacks "sing" long (half-hour) and complex "songs", apparently as part
of lek behavior. also called arena behavior.
Lek behavior is a kind of courtship exhibited by many species of insects, birds and mammals. A number of males gather in a defined area, each in his own piece of the space, and engage in competitive mating displays. Females cruise through the arena, observe the displays, and choose a mate based on their evaluation.
In the case of the humpbacks, the male display appears to be a song. Humpback songs have been recorded (by Naval sonar engineers) since the WW II era. As a result, the following curious pattern has been observed. At any given time, all the humpbacks in a given region are singing basically the same song. Individual performances may differ in small ways -- an extra trill here, a note left out there, a section repeated somewhere else -- but the basic pattern is the same. Over time, the pattern evolves, until after a year or so, it is completely changed. All the males are still singing the same song, but it is a completely different song than it was before.
After about 50 years of recordings, the old songs have never come back into fashion. The evolution appears to be open-ended.
Humpbacks around Bermuda sing completely different songs from humpbacks around Hawaii, even though they seem to be entirely the same species.
As far as anyone can tell, the patterns in the songs have no meaning at all. They are just abstract patterns. Pure style, no content -- in this sense, at least, like human melodies.
It remains unclear to what extent the development of the song is a matter of social construction -- everyone following a gradually shifting fashion -- and to what extent it is implicit in a given stage of the song -- that is, the "next thing" is obvious to any humpback with an ounce of style.
In any case, it is tempting to analyze this phenomenon by reference to the human phenomenon of fashion. You can -- and must -- distinguish yourself by a slightly individual interpretation of the current style, but last season's stuff is totally lame, and so is some random set of choices that indicate you're completely out of touch with the culture.
Grammar -- without communication
There are some forms of complex behavior that do not really seem to be properly viewed as displays, since they are done just as often by solitary animals, conspecifics seem to pay no attention to them, etc. Some such behaviors nevertheless seem to have a complex, quasi-grammatical structure.
The best-studied case is mouse grooming.
Mice use their forepaws to spread secretions from oil glands near their eyes. They do this via a rapid series of motions, which may be forehand or backhand, up or down, from the midline to the side or vice versa, etc. The particular order of such gestures has no functional consequence, as long as there are enough of each type to spread the secretions around.
Any given mouse uses a stereotyped sequence of such gestures -- which can be appropriately described with a finite-state grammar -- but different mice use different sequences. Genetically identical mice use identical sequence patterns.
Thus this is a case where a grammar-like organization of behavior is found, though there seems to be no communicative significance.
The organization of humpback whale songs is different in some ways -- it changes rather than being genetically fixed, and other whale seem to be paying attention to its structure. However, whale song syntax is similar to mouse grooming syntax, in that (as far as we know!) the structure does not seem to have any significance other than itself.
A caveat
We might be -- we almost certainly are -- missing a lot of the subtleties of animal communication, despite years of careful and patient observation by ethologists.
Size of the repertoire
It is generally said that individual animals may have a repertoire of up to about 40 different displays. One authority says:-
For most relatively social adult fishes, birds and
mammals, the range or repertoire size for different species varies from
15 to 35 displays.
Curiously, there is little correlation with location in the "great chain of being." Cuttlefish, as far as we know, have about the same repertoire size as non-human primates do.
The hypothesis advanced about this constancy of repertoire size is that there is a sort of dynamic equilibrium involved -- old displays tend to be lost as new ones are added. Presumably there are some pressures to prevent the accumulation of displays: they may become ecologically irrelevant; there may be too much investment required in the brain circuitry involved; it may be confusing to have too many "lexical items" in the system, lessening the effectiveness of each.
One clear evolutionary puzzle is why -- if communication systems are as useful as we generally think they are -- the rest of the animal kingdom seems to have put so little emphasis on elaborating them beyond this size of a few tens of hard-wired elements.
How human language seems to be different
You should be able to fill this section in for yourself.
The basic issues are the phonological principle and the elaboration of a large (and variable) lexicon; and the existence of a hierarchical compositional semantics, where the meaning of the whole is a function of the meaning of the parts in some way more complex than simple summation of the correlates of independent displays.
In addition, human communication -- by means of spoken language or in other ways -- seems to involve a much better-developed "theory of mind", involving more conplex and abstract models of others' knowledge, beliefs, intentions and goals, and much more complex and systematic use of these models to plan sequences of communicative acts. Indeed, it remains a matter of debate whether other animals can be said to have a "theory of mind" at all, as opposed to innate automatisms (like gaze following) or learned reactions that make it seem in some cases as if they were communicating in the same way.
At the same time, human facial expression, (aspects of) human "body language", and (aspects of) human voice modulation in speaking, seem quite similar in kind to animal displays, and indeed similar in detail in many cases to the expressions and gestures of other primates.
Whether one chooses to emphasize the continuities or the divergences is to some extent a matter of taste. However, those whose interest is the form and meaning of human languages do not, in most cases, find very many points of continuity with the communicative practices of non-human animals.